Как оптимизация калийного питания помогает подавить развитие соевой тли
Соевая тля (Aphis glycines Matsumura) – инвазивный вид насекомых, выявленный в США в 2000 г. Осмотр полей и наблюдения в штатах Висконсин и Мичиган (США) показали, что на многих полях из числа наиболее сильно зараженных соевой тлей у растений сои проявлялись также внешние признаки недостатка калия. В данной статье обобщены результаты недавних исследований по изучению взаимосвязей между уровнем калийного питания сои и интенсивностью заселения растений тлей. Целью исследований было установить, как оптимизация минерального питания растений помогает в борьбе с этим вредителем.
Штат Висконсин, 2001-2002 гг.
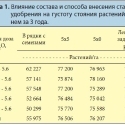
В мелкоделяночном полевом опыте с внесением калийного удобрения в разных дозах содержание калия в листьях сои и урожайность повышались с ростом содержания подвижного калия в почве (табл. 1), но статистически значимых различий в численности тли между вариантами опыта выявлено не было (Myers et al., 2005). Проводившиеся опрыскивания инсектицидом снижали численность тли и повышали урожайность сои, но статистически значимого взаимного влияния обработок инсектицидом и питания растений калием на данные показатели выявлено не было.
При этом численность тли и в первый, и во второй год указанного опыта была очень высокой – существенно выше, чем на полях фермеров. Например, в 2002 г. пиковая численность тли на необработанных инсектицидом делянках превысила 1600 особей/растение, а средняя пиковая численность данного вредителя, выявленная при обследовании полей сои в южной части штата Висконсин, составила 280 особей/растение. Возможно, из-за близкого расположения (<0.9 м) и небольшого размера делянок (3.0 х 7.0 м) растения, испытывавшие сильный недостаток калия, привлекали и служили источником пищи для больших популяций тли, что приводило к колонизации и достаточно обеспеченных калием растений. Таким образом, примененная разбивка делянок в данном полевом опыте могла помешать выявить взаимосвязи, которые, по-видимому, существуют в масштабах всего поля.
Штат Висконсин, 2003 г.
В 2003 г. в специальном лабораторном опыте была изучена плодовитость тли при питании на листьях сои, отобранных на опытном поле в Арлингтоне (штат Висконсин) со здоровых растений и растений с внешними признаками недостатка калия. Количество личинок, приходящихся на одну взрослую особь, и скорость роста популяции были существенно выше при питании тли на листьях сои с низким содержанием калия (табл. 2). Подобное действие калия свидетельствует о том, что его недостаток у растений сои потенциально способствуют более высокой скорости роста популяции тли. Однако в контролируемых лабораторных условиях исключается действие такого фактора, как естественные враги тли (хищники и паразиты), которые существуют в полевых условиях.
Механизм действия калия в вышеуказанном опыте не изучался, но, как отмечают другие исследователи, пищевой рацион тли формируется за счет растворимых аминокислот, а недостаток калия может вызывать увеличение концентрации этих аминокислот в растительных тканях.
Штат Висконсин, 2004 г.
В штате Висконсин в 2004 г., когда натиск вредителей был несильным, был проведен мониторинг численности соевой тли в производственных посевах (34 поля) при диапазоне содержания подвижного калия в почве от 80 до 200 мг К/кг почвы и выше (Myers and Gratton, 2006). Встречались и поля с почвами песчаного гранулометрического состава, а для них критическое содержание подвижного калия (верхний предел “низкого” класса обеспеченности) составляет уже 60 мг К/кг почвы. Скорость роста популяции тли на полях с песчаными почвами отрицательно коррелировала с содержанием подвижных форм K и P в почве, а также с содержанием K, N, P и S в листьях растений. Однако пиковая плотность заселения растений тлей положительно коррелировала со всеми вышеуказанными почвенными и растительными показателями.
Проведенный в том же году полевой опыт по изучению действия разных уровней калийного питания на развитие тли показал, что при среднем классе обеспеченности почвы подвижным калием и выше снижалась скорость репродукции тли, замедлялась скорость роста популяции и уменьшалась пиковая численность естественной популяции тли (табл. 3). К листьям растений, незаселенных тлей, с помощью зажимов прикреплялись маленькие садки, что позволило в полевых условиях изучить численность потомства одной особи тли. Тля помещалась на лист сои в небольшое ограниченное пространство, изолированное от других тлей и защищенное от хищников.
Не ясны причины того, почему в 2001 и 2002 гг. численность тли не снизилась при более высоком содержании подвижного калия в почве (табл. 1), как это произошло в 2004 г (табл. 3). Данное обстоятельство может быть связано с меньшим натиском вредителей в 2004 г., что и позволило выявить влияние минерального питания растений на развитие тли. Высокая же численность тли, наблюдавшаяся в 2001-2002 гг., не дала этого сделать. При этом размер делянок в 2004 г. был таким же, как и в более ранних опытах.
Штат Мичиган, 2003-2004 гг.
В юго-западной части штата Мичиган в середине августа 2003 и 2004 годов были обследованы производственные посевы сои (5-8 полей) с выявленными внешними признаками недостатка калия у растений (Walter and DiFonzo, 2007). В пределах каждого поля выделялось 2 участка: один выбирался в центре области с внешними признаками сильного недостатка калия у растений, а другой – в ближайшей части поля, где растения не проявляли признаков недостатка калия. На каждом из этих участков отбирались образцы почвы, проводился сбор флоэмного сока растений, а также определялась численность тли. И в первый, и во второй год обследований содержание обменного калия в почве было ниже на тех участках полей, где у растений сои проявлялись внешние признаки недостатка калия. В 2003 г. произошла вспышка численности вредителя, и плотность заселения растений тлей была выше также на участках полей, где наблюдались внешние признаки недостатка калия у растений (табл. 4). Численность тли в 2004 г. была крайне низка для того, чтобы выявить различия в плотности заселения растений этим вредителем.
В округе Ван-Бюрен (штат Мичиган) в 2004 г. был проведен полевой опыт в производственных посевах сои с низким содержанием обменного калия в почве. Исходное содержание обменного калия в почве составило 67 мг К/кг почвы, что свидетельствует о недостаточной обеспеченности почвы калием. Критический уровень для данной почвы (с ЕКО, равной 8.6 мг∙экв/100 г) составляет 96 мг К/кг почвы. В опыте было два варианта: без внесения и с внесением калийных удобрений в дозе 157 кг K2O/га. Опыт проводился в 5-ти кратной повторности, площадь делянки составила 6.1 х 36.6 м. Для наблюдения за репродуктивной способностью тли использовались маленькие садки и большие полевые садки. Маленькие садки, где в небольшом ограниченном пространстве тля была защищена от хищников, прикреплялись к листьям сои с помощью зажимов (как указывалось выше). Данные садки применялись для изучения роста и численности потомства одной особи тли. Большие садки представляли собой куб со стороной 1 м, и ими накрывалось по 10 растений. Данные садки также защищали помещенных внутрь тлей от хищников, но, в отличие от мелких садков, позволяли улетать появляющимся крылатым (летающим) особям.
В маленьких садках, установленных в первый раз – 10-го июня, различий в репродуктивной способности тли между вариантами опыта выявлено не было. В аналогичных садках, установленных во второй раз – 14-го июля, личинки тли появлялись раньше и в бóльших количествах на растениях сои, не получавших калийного удобрения, по сравнению с удобренными калием растениями (табл. 5). В больших садках заселение тлей проводилось 28-го мая из расчета одна особь/растение. Начиная с 30-го июня, в больших садках наблюдалось статистически значимое увеличение численности тли в варианте без внесения калийного удобрения.
Во всех опытах, проводившихся в Мичигане в 2003 и 2004 годах, были проанализированы образцы флоэмного сока растений. При этом определялось соотношение 18-ти незаменимых аминокислот без измерения их общего содержания во флоэмном соке. Была установлена обратная корреляция между относительной долей аминокислоты аспарагина и содержанием обменного калия в почве, в то время как для других аминокислот подобных зависимостей выявлено не было. Таким образом, содержание аспарагина в растительном соке повышалось при снижении содержания обменного калия в почве: при содержании обменного калия, равном 120 мг К/кг почвы, на долю аспарагина приходилось 3-10% от суммы аминокислот, а при 20 мг К/кг почвы доля аспарагина возрастала до 8-20%.
Аспарагин может играть критическую роль в питании тли азотом, поскольку тля обладает ограниченной способностью усваивать азотсодержащие соединения. Вейбулл (Weibull, 1988) отмечал, что растительный сок наиболее устойчивых к тле образцов овса и ячменя содержит относительно мало аспарагина. Как указывали Ричардс и Бернер (Richards and Berner, 1954), недостаток калия вызывает повышение содержания аспарагина в листьях ячменя. Баркер и Брэдфилд (Barker and Bradfield, 1963) сообщали, что увеличение концентрации калия в питательном растворе приводит к снижению содержания свободных аминокислот, особенно аспарагина, в молодых проростках кукурузы.
Считается, что тля получает весь свой рацион азота за счет аминокислот, транспортируемых с флоэмным соком растений. Как известно, в процессе пищеварения у тли не используются протеиназы, возможно потому, что во флоэмном соке обычно высоко содержание ингибиторов протеиназ и крайне низка концентрация белков. По этой причине растительные белки являются плохим источником азота для тли. По сообщению Годфрея и Хатчмахера (Godfrey and Hutchmacher, 1999), внесение калия под хлопчатник в штате Калифорния (США) в дозах от 112 до 224 кг K2O/га оказывало “умеренное отрицательное влияние как на срок развития генерации тли, так и на ее плодовитость.” Таким образом, по мере усиления стресса, вызванного недостатком доступного калия в почве, растения реагируют выделением во флоэму большего количества свободных аминокислот, таких как аспарагин. Это делается для того, чтобы уравновесить осмотический дисбаланс в растениях. Тля же использует эти свободно перемещаемые и легкоусваиваемые азотсодержащие соединения с выгодой для себя – она быстрее развивается и дает больше потомства в расчете на одну самку. Это приводит к более быстрому росту численности тли и, в конечном итоге, к более высокой плотности заселения растений тлей, что увеличивает потери урожая.
Выводы
С учетом содержания стандартно определяемых подвижных и доступных форм калия в почвах штатов Висконсин и Мичиган повышение численности тли на соевых полях наблюдалось только в тех случаях, когда содержание данных форм калия опускалось до минимальных значений, и было значительно ниже уровней, рекомендованных при возделывании сои. Почвенное обследование, проведенное в 2005 г., показало, что медианные значения содержания обменного калия в вышеуказанных штатах составили, соответственно, 125 и 149 мг К/кг почвы. При этом доля почв с содержанием обменного калия ниже 80 мг К/кг почвы оценивалась примерно в 10-15%.
Хотя результаты, полученные в штатах Висконсин и Мичиган, и свидетельствуют о сильном влиянии на развитии соевой тли такого фактора, как питание растений сои калием, это отнюдь не означает, что достаточная обеспеченность растений этим элементом питания может гарантировать стопроцентную защиту от соевой тли. Численность тли контролируется еще и естественными врагами, такими как хищная коровка хармония (Harmonia axyridis Pall.), а также паразитами. Это примеры трофических факторов, подавляющее действие которых на тлей проявляется с разной силой в зависимости от условий конкретного года и региона, в сравнении с факторами, связанными с минеральным питанием растения-хозяина. Заселение растений тлей может происходить и при достаточном уровне калийного питания растений.
Тем не менее, предотвращение недостатка калия у растений – это, по крайней мере, одна из мер защиты от тли и страховка от потери урожая из-за повреждения этим потенциально опасным вредителем, которое к тому же является и переносчиком вирусных и грибных болезней растений. С практической точки зрения это означает, что при возделывании сои необходимо поддерживать плодородие почвы по калию на рекомендуемом уровне, поскольку это является составной частью интегрированной системы защиты растений от соевой тли.
Д-р Граттон – адъюнкт-профессор кафедры энтомологии Университета Висконсина (г. Мадисон, США). Д-р Ди Фонзо – профессор кафедры энтомологии Университета штата Мичиган (г. Ист-Лансинг, США). Д-р Бруулсема – Директор Международного института питания растений по Северо-Восточному региону Северной Америки (г. Гуэльф, провинция Онтарио, Канада); e-mail: tom.bruulsema@ipni.net.
Перевод с английского и адаптация: В.В. Носов.
Редакция: С.Е. Иванова
Литература
Barker, A.V. and R. Bradfield. 1963. Agron. J. 55(5): 465-470
Godfrey, L.D. and R. Hutchmacher. 1999. http://www.cdfa.ca.gov/is/fflders/ completed_projects.html
Myers, S.W., C. Gratton, R.P. Wolkowski, D.B. Hogg, and J.L. Wedberg. 2005. J. Econ. Entomol. 98: 113-120.
Myers, S.W. and C. Gratton. 2006. Environ. Entomol. 35: 219-227.
Richards, F.J. and E. Berner, Jr. 1954. Annals of Botany 18: 15-33.
Walter, A.J. and C.D. DiFonzo. 2007. Environ. Entomol. 36(1)26-33.
Weibull, J. 1988. Phytochemistry 27: 2069-2072.
Скачать материалы:
Как оптимизация калийного питания помогает подавить развитие соевой тлиSize: 1,64 MB